
Site Map
Week 1 Lecture Notes
Terminology
As you go through the readings, especially in journal articles, you will likely notice that there seems to be a lack of agreement on terminology within the field of navigation and spatial representation. The simple fact is that different groups of researchers use the same terms to mean wildly different things. As you might imagine, this tends to complicate matters, especially if you are interested in interdisciplinary or cross-species comparisons. Hopefully, these notes will help give you at least a little sense of how the terminology has developed.
Since Aristotle people have wondered (and written) if some birds, but not others, migrate. Or, did the birds transmute into other species, hibernate in mud, or fly to the moon? Aristotle opted for the migration explanation, but it wasn't until the end of the 19th century that this was a generally accepted fact.
Of course, some birds do not head for warmer areas in the winter. This seemed to divide birds quite nicely into two categories: migratory and non-migratory (i.e., resident) birds.
Heape (Emigration, migration and nomadism, 1931, Cambridge: Cambridge University Press) tried to find other animals that migrate like birds (i.e., were "true migrants"). At this point in time some animals, including some fish and whales, were accepted as migratory. The rest were, like resident birds, classed as non-migratory. Heape noticed that some animals that were classed as non-migrant showed intriguing movements. This resulted in a required further sub-division of the terminology. Heape proposed the terms nomadism and emigration.
Nomadism described animals that appeared to wander about without returning consistently to the same location at the same time each year. Emigration implied movement away from unsuitable conditions. It was soon noticed that such movement usually produced colonizers as well as refugees. For the latter, the term emigration was inadequate. Subsequently, the term dispersal, was adopted because emigration had usually been applied to situations in which animals scattered in all directions from their place of origin. Unfortunately, as time would demonstrate, this was an even worse choice of terminology that the inadequate term emigration.
Mammologists were the first to adopt the term dispersal wholeheartedly. They used the word to describe the behaviour of young mammals that abandoned the area they were born in to settle elsewhere. Ornithologists adopted the term dispersal for a similar behaviour: the movement of just fledged birds away from the nest. However, the term quickly came to be used by both mammologists and ornithologists to describe changes in breeding sites which resulted in the segregation of previously cohesive breeding groups. By the 1960s all non-trivial movement was either described as migration, dispersal, or nomadism. In other words, anything that wasn't migration or nomadism was dispersal. The original meaning of the dispersal was quickly forgotten and was subsequently used in a variety of semantic corruptions.
The verb "scatter" means "to disperse". It applies to groups of things leaving an area. However, it was quickly, and inappropriately, applied to individuals. The only way an individual can, technically, scatter is...messy.
Entomologists of the 1950s perceived insect migration as directed and involving back and forth seasonal reversals (e.g., the seasonal migration of Monarch butterflies). In the late 1960s an argument arose in the entomological community against directed insect movements; what had previously been defined as migration was now termed dispersal by the school of thought championed by C.G Johnson (1969). In fact, all non-trivial insect movements were now called dispersal, with a total disregard for Heape's original definition. The final corruption came when the convergence of some insects upon hibernation sites in vast numbers was included under the general term of dispersal.
Johnson subsequently redefined migration and dispersal as being synonymous, calling migration "the adaptive change of breeding habitat". Of course, this was pretty much the same definition mammologists and ornithologists used for the term dispersal! By 1970 mammologists and ornithologists defined migration as the to-and-fro return movement that ended where it began (i.e., at the original breeding site). Entomologists used the term in exactly the opposite sense: to indicate on change in breeding sites.
Completely segregated from this discussion where the demographers, anthropologists, and social scientists who were independently attempting to develop their own terminology. Quite by chance they ended up allying themselves with entomologists, rather than with those groups who studied other vertebrate groups. Researchers dealing with human movement tended to use migration and dispersal interchangeably, but the emphasis was slightly different from how entomologists used the terms. While entomologists said migration was really dispersal, human demographers said that dispersal is really migration. As a final twist, humans that performed seasonal return migrations, comparable to similar feats performed by birds, were not classifed as migrants, but as nomads!
Top
Schools of Thought
Broadly speaking, four schools of thought, or methodologies, are commonly seen to guide much of the navigation research in the field of psychology. Traditionally, researchers of navigation took their guidance from ethology, behaviourism, or behavioural ecology and sociobiology (or some combination thereof). More recently cognitive neuroscience is becoming increasingly important to researchers of navigation and spatial representation.
Ethologists (e.g., Tinbergen, Lorenz, von Frisch) are interested in animal behaviour, typically behaviour that occurs in an animal's natural environment. Ethology was developed by European students of animal behaviour. Originally, this school of thought viewed all behaviour in terms of fixed action patterns and sign stimuli. Ethologists argued that migration patterns of animals are pre-wired, genetically controlled programs that have evolved to allow animals to survive. The animals, it was argued, have little or no control over where or when they go someplace. Some learning was permissible (e.g., imprinting), but it was immediate or irreversible. These factors can be influenced by environmental features and, perhaps, the animals have some small say in which clearing to rest in, but by and large, the animals were treated as automatons. The early ethology perspective just described has shifted somewhat in modern ethology, although there is still a certain predisposition to look for hard-wired, genetically regulated, behaviours. Many modern ethologists are more interested in the behaviour that results from the complex interplay between an animal's behavioural predispositions and environmental features.
Behaviourists, championed by American psychologists like Watson and Skinner, maintained that animals migrate/navigate through a series of learned behaviours. Internal programs or drives were considered irrelevant. It was the early learning patterns that were seen as being extremely influential in determining how animals migrate. Observational learning is seen to play a major role, especially the observation of parents or other experienced adults by juveniles. The radical behaviourist approach (e.g., Watson and Skinner's theories that argue against making any inference judgements about internal drives) waned with the cognitive revolution of the 1970s. Nevertheless, there is still a great deal of merit in considering the impact of observational learning and reinforcement schedules when considering many questions of short and medium range navigation (this is especially true for certain social birds and mammal species).
Behavioural ecologists (e.g., R. Robin Baker) and sociobiologists (e.g., E.O. Wilson) are a third group of researchers who have recently become invovled in the study of navigation and migration. Behavioural ecology truly come into its own through the late 1970s and early 1980s. Generally, behavioural ecologists are interested in animal behaviours that serve to increase the reproductive fitness of a species. However, at the level of the individual behavioural ecologists maintain that animals are individuals confronted with dangers and problems that must be sovled if they are to survive and reproduce. Behavioural ecologists agree that animals come equipped with genetic predispositions (as argued for by ethologists) and are more than capable of learning through conditioning (as argued for by behaviourists). Social scientists argue that man is a sentient creature. Behavioural ecologists suggest that there is no good reason for assuming this feature is unique to humans, and maintain that all animals are, to greater or lesser degree, sentient. Animals are individuals, packed with predispositions and learned experiences. They are the produce of natural selection and are sentient creatures prepared to solve any problem a complex environment can throw at them, or die in the attempt.
Cognitive neuroscience is the most recent methodology to leave its mark on the study of navigation. Generally speaking, cognitive neuroscientists are interested in how underlying neural mechanisms and organization of the central nervous system guide and control organisms' thought processes and behaviours. As such, many of the new generation of psychologists may be very broadly classified within this category. Cognitive neuroscientists often make heavy use of number of technological resources, such as fMRI, lesioning studies, and drug delivery to brain regions through permanent canulas. On the one hand, this allows for a greater understanding of the involvement of brain mechanisms in navigation and spatial representation. On the other hand, the technology often requires that the research be conducted in laboratory settings, resulting in some limitations in movement ranges. For questions about short-scale navigation, such as foraging or choice behaviours, this isn't much of a problem. However, it is currently difficult (or expensive) to study large-scale navigation, such as seasonal migration, using this methodology.
Top
Kinesis and Taxis
Kinesis involves movement in a random direction. A stimulus (e.g., light, moisture, temperature) is presented and the animal begins moving. When the stimulus is removed the animal stops moving. It is important to remember that kinesis is not directed movement. That is, the animal isn't moving towards or away from something, it is just moving. It will continue moving until the motivating stimulus is no longer present.
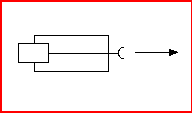
Figure 1: A schematic representation of an agent with a source of locomotion at the rear and a receptor at the front. Stimulation of the receptor, which directly controls the motor response, causes the agent to move forwards.
Taxis involves directed movement. That is, movement that bears some relationship to the location of a stimulus. In this case, in the presence of a stimulus the animal proceeds to move either towards or away from the source of the stimulus. For example, the larval stage of Chaoborous, a small midge, is aquatic. The larvae demonstrate a daily verticle movements up and down in the water column due primarily to light levels: In the presence of light they move up through the water column, without light they move down.
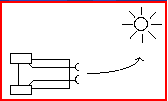
Figure 2: A schematic representation of an agent with two motor control systems, allowing purposeful turning. In this case, two lateralized sensors each connect to the same-side motor system. In this arrangement, stimulation of the left receptor, for example, will activate the left drive system, causing the agent to turn to the right, and vise versa. This arrangement would produce an organism that moves away from, for example, a light source. If a slight alteration was made, and the sensors were attatched to the opposite motor system (e.g., left sensor to right motor, and vise versa) the agent would, instead, move towards the stimulus.
Depending on who you talk to, tropisms may or may not be viewed as simple forms of navigation. I am inclined to call tropisms orientation responses. As such, they may be considered the minimum requirement for a navigating organism.
Top